Subtext goes right here whatever you want to say goes here
All rights are reserved. Please cite this page for non-commercial use. Contact us for commercial use.
Citation: Cliff Shunsheng Han, Oral Probiotic Deficiency May Cause Common Allergies – Theory of Negative Trigger Marks the Interaction between Microbiota and Host Immune System. Website: https://allerpops.com/oral-probiotic-deficiency-may-cause-common-allergies/, published by AllerPops Corp on 11/17/2019.
Cliff Shunsheng Han
1 Knoze Jr Corp., 1650 Trinity Dr., Suite 103, Los Alamos, NM 87544.
*Correspondence to Cliff S. Han, Knoze Jr Corp., 1650 Trinity Dr., Suite 103, Los Alamos, NM 87544. Email: cl******@*******ps.com; 505-695-4236
This study seeks to identify causes for common allergies and to develop translational measures to treat common allergies such as allergic rhinitis. For this reason, I performed a longitudinal, cross-sectional, and translational study of oral microbiota that lasted for three years. With the help of an unusual allergy case of an immigrant, I identified the cause of his allergies. The attempt to correct the cause was a great success with lasting allergy relief.
The study results indicate that the direct mechanism behind common allergies may be an oral probiotic deficiency. In other words, the lack of beneficial bacteria, Streptococcus, and Veilonella, in the oral/nasal cavities causes allergies. These bacteria are mutualists. Together, they produce metabolites that pacify the nearby immune system. With this in mind, I developed a prebiotic mix to promote those probiotics. As a result, the subject reached long-term remission from his allergies.
In addition, the study led to the development of the Theory of the Negative Trigger (TNT). TNT advocates that oral probiotics are a negative trigger controlling the immune system, so the host and hosted microbiota can make peace with each other. As a consequence of the theory, we may modulate the interaction between microbiota and the immune system to prevent, treat, and even cure allergies, autoimmune diseases, and potentially cancers.
List of abbreviations: TNT, Theory of the Negative Trigger; OTU, operational taxonomic unit; QIIME, Quantitative Insights Into Microbial Ecology
More than 30 years ago, Dr. Strachan proposed the hygiene hypothesis to explain the increase in allergy diseases (Strachan, 1989). Subsequently, many studies suggest that personal and social hygiene practices are associated with the allergy disease epidemic (von Mutius, 2007). The hypothesis currently emphasizes the exposure of diverse environmental bacteria rather than infection (Ege et al., 2012; Jatzlauk et al., 2017). Allergic subjects are found to have microbiota dysbiosis (Chung, 2017). Indeed, certain bacteria interact with the immune system through metabolites or structural molecules produced by bacteria (Blacher et al., 2017; Gensollen et al., 2016). However, attempts at using probiotics to cure or prevent allergy diseases have had limited success (West et al., 2016).
In all those trials, researchers used probiotics that live in the gut. Instead of focusing on the remote interaction between the gut and the airway, I examined the local interaction between microbiota living in the airway and the immune system surrounding the respiratory tracts. As a result, this study illustrates that restructuring oral microbiota can lead to lasting remissions of common allergies (allergic rhinitis).
This article uses “allergies” when referring to common allergies and allergic rhinitis.
The subject (CSH, the author) started having allergies in the spring of 2014, eighteen years after immigrating to the US in the late nineties. His symptoms included nasal congestion, rhinorrhea, red and itchy eyes, tearing, coughing, aches in the throat, and a tight chest. According to a study from Italy, immigrants usually develop common allergies within three to five years after immigration (Lombardi et al., 2008). Unlike those typical cases, this eighteen-year lapse between immigration and the development of the symptoms is extremely long. This long lapse might indicate that the underlying cause was likely to be personal rather than environmental.
Prior to the appearance of the first symptoms, the subject intensified his oral hygiene routine. In his first ten years in the US, he only brushed his teeth once daily. Gradually, he began introducing additional dental hygienic practices, such as dental cleaning, flossing, tongue scraping, and using mouthwash. Eventually, before allergy symptoms first appeared in the winter, CSH brushed and flossed his teeth once daily. He also scraped his tongue several times a week and used mouthwash frequently. In addition, he took antibiotics for a week to treat a lasting cough. Researchers have observed the association between the use of antibiotics and the subsequent appearance of allergic symptoms (Foliaki et al., 2009).
After reflecting on his life in the past 18 years, CSH suspected that changes in his oral microbiota might have contributed to the development of his allergies. So, he began to collect samples of his saliva and, for comparison, fecal matter periodically. Altogether, the sample collection continued until spring 2017 (Table 1). However, I present only data from saliva samples here, as no positive result was found from fecal samples.
The first episode of CSH’s allergic rhinitis lasted a year and a half. It ended in the summer of 2015 and became seasonal afterward. In other words, he partially grew out of his allergies. During this time, CSH experimented by reducing his oral hygiene for several weeks. Consequently, local moderate periodontitis and bleeding, and gum recession developed, but his allergy symptoms persisted. Resuming brushing and flossing for two weeks healed the inflammation, while the allergy symptoms continued. After that time, his only oral hygiene practice was brushing his teeth daily without toothpaste. His allergy symptoms were alleviated during two four to eight-week trips abroad but never completely disappeared before the summer of 2015 (Timeline in Table 1 and Figure 1A).
For comparison, saliva samples of his family members were also collected (Table 1). The samples collected before 2017 were sequenced and analyzed using a commercial service (Supplemental material). Yet, the result showed no significant difference between the fecal samples before and after summer 2015 (data not shown).
The results of CSH’s saliva samples show that the relative abundance of Veillonella, a helpful bacteria, increased (p=0.042) in saliva samples collected after the summer of 2015 (Figures 1A, 1B). Besides that, an increase in Streptococcus was not significant. The initial analysis indicated that the difference in abundance of Veillonella in CSH and his family members is insignificant. However, one family member also suffers from seasonal allergies. His saliva had Veillonella at half of those of non-allergic family members. The average abundance of Veillonella in the two non-allergic family members was five to six times as high as in the two subjects with allergies. Also, two of the three OTUs (operational taxonomic units) in the genus of Streptococcus had similar distribution patterns in the family as Veillonella (Figure 1C).
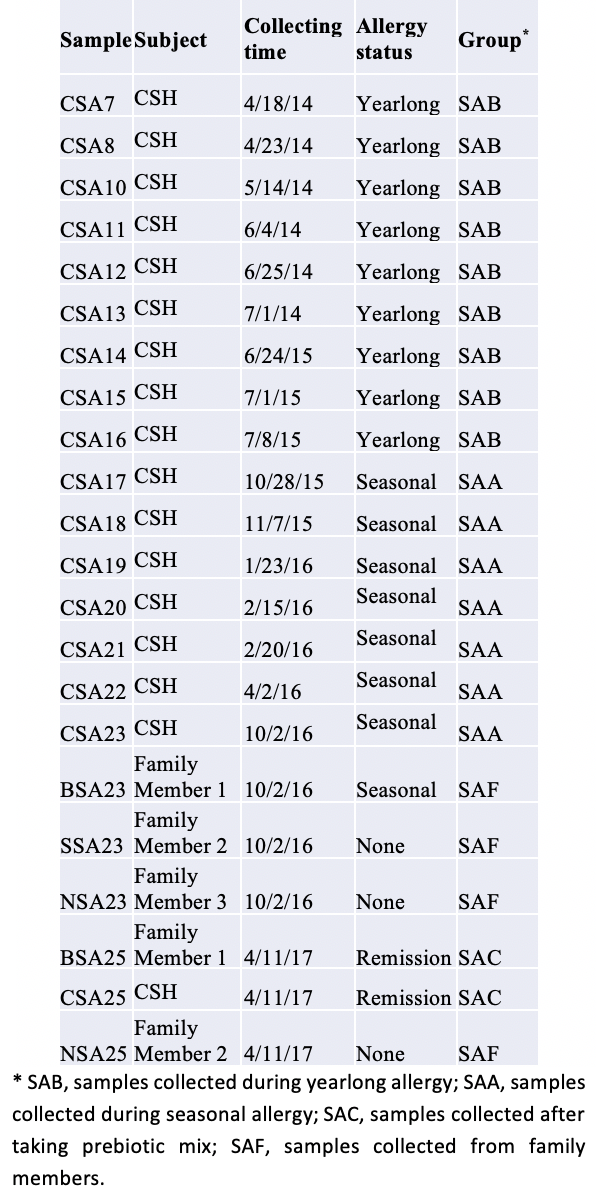
Numerous studies indicate that Streptococcus and Veillonella grow together in the oral cavity with a mutualistic relationship (Takei et al., 1968; van der Hoeven et al., 1978). Streptococcus attaches itself to surfaces, and Veillonella metabolizes the cavity-inducing lactic acid from the Streptococcus and converts it into short-chain fatty acids (SCFA). SCFA are less toxic than lactic acid and benefit the human host in many other ways. One is especially relevant to this study: they interact with the immune system to reduce inflammation (Correa-Oliveira et al., 2016).
Like the gut, the respiratory tract is another vulnerable interface between humans and the outside world. The immune system, such as the tonsil, surrounds the airway from the mouth/nose to the bronchi. Because the results showed reduced probiotics when the allergies were severe, I formed a hypothesis: not having enough oral probiotics is the cause of allergic rhinitis. In other words, we may moderate or eliminate allergy symptoms if we use prebiotics to raise the level of relevant, beneficial bacteria in the oral cavity.
Given these findings, I developed a prebiotic mix (U.S.Pat. No. 9,795,579). The composition contains sugars and arginine. The ingredients promote the growth of selected bacteria. They are picked based on the fermentation profile of Streptococcus and Veillonella, both as individual species and as a community (Han, 2017; Kolderman et al., 2015; Willcox, 1996).
Both CSH and the other family member with allergies took the prebiotic mix several times a day in the early/middle of March 2017 when their allergies were at their peak. Those attempts alleviated the allergy symptoms significantly. Neither subject had to take anything else for their allergies. However, the relief from their symptoms was temporary. They had to take the oral prebiotic mix several times a day to have continued success.
Five days after taking the prebiotic composition, CSH developed a fever, likely due to a bronchi infection. He possibly contracted the infection from another family member who had become sick a week earlier. His temperature was between 38-39०C(100-102०F), accompanied by a phlegmy cough. CSH stopped using the compound once the fever began.
On the second day of the illness, he noticed his tongue was completely red and missing its usual biofilm. Like a farmer facing a burned field, he suspected that his fever/infection had cleared the microecospace in his oral cavity. The fever produced a unique opportunity for restructuring his oral microbiota.
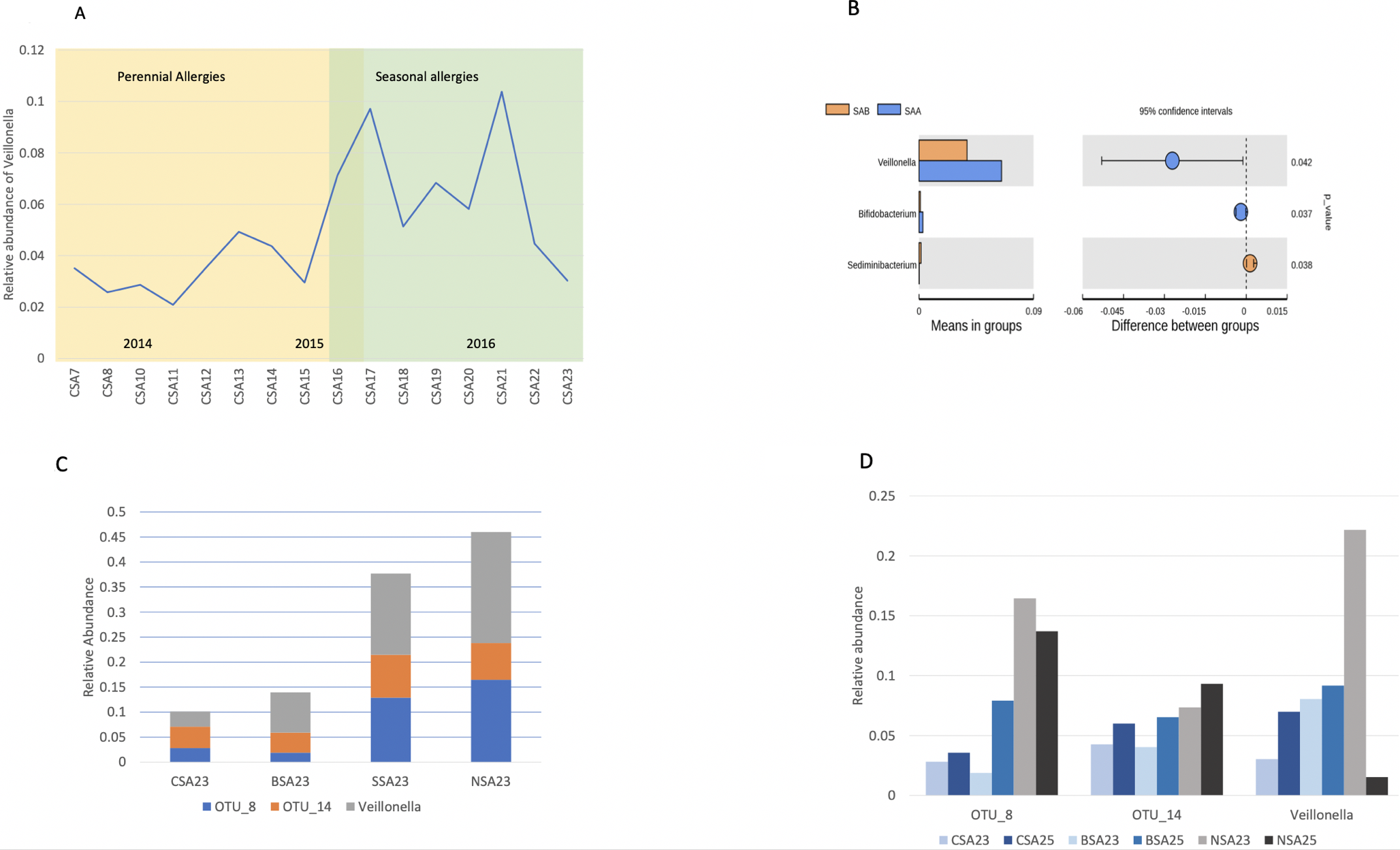
CSH took two doses of the prebiotic mix in response when his fever receded. Surprisingly, he found his allergic symptoms gone the next day, March 15, 2017. He developed a hot-water-based method to clear the oral microecospace for the other family members with allergies (Supplementary Figure 1). That was scraping the tongue with a wet washcloth. Then, heat the tongue and gums by gargling and swishing hot water for more than five minutes. This allows the prebiotics better access into the body. The family member took some of the prebiotic mixes immediately after cleaning. His allergies went into remission as well. After that, CSH and this family member no longer experienced allergies for the rest of the 2017 spring season.
The results indicate that our hypothesis is correct: oral probiotic deficiency is likely the underlying cause of allergies. To further confirm the theory, I wanted to see if the targeted bacteria increased after the treatments.
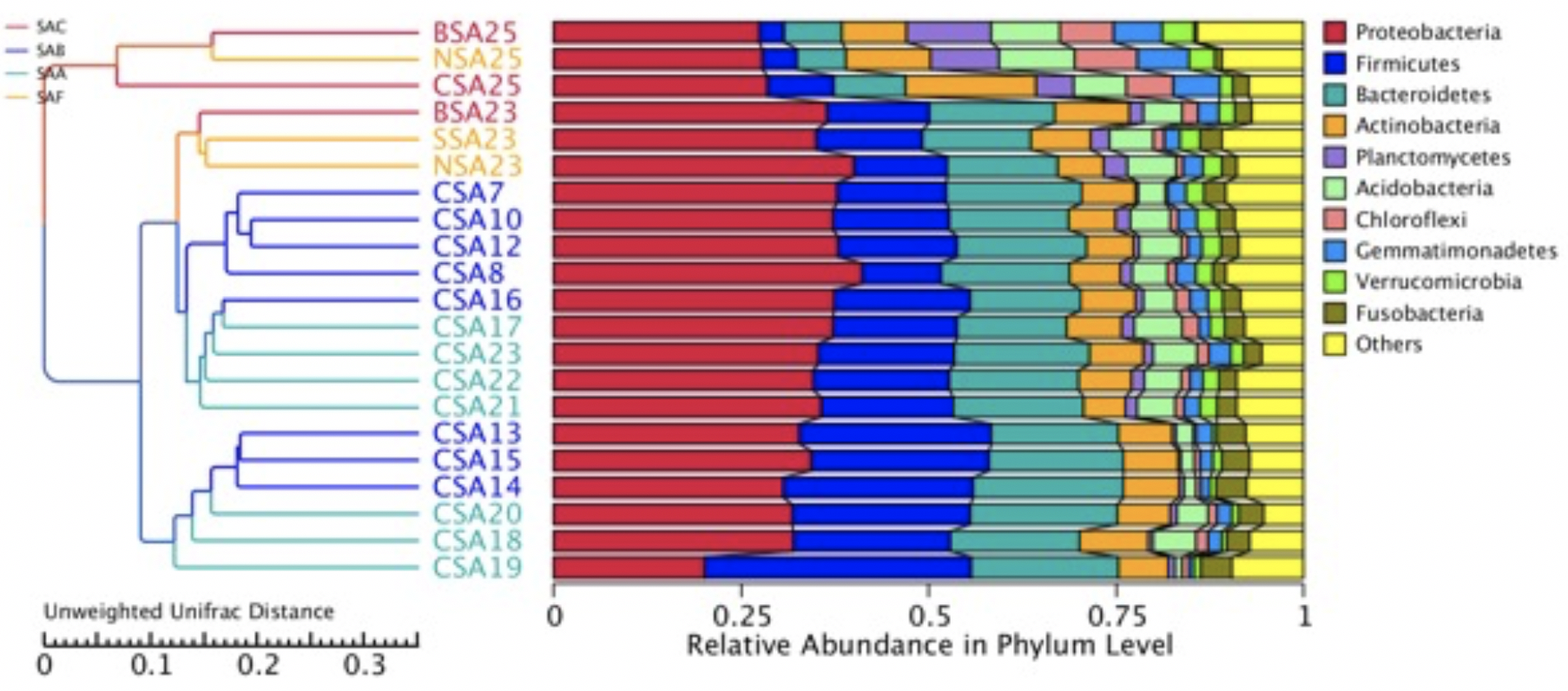
Approximately three weeks after their remission, the last batch of saliva samples was collected. I sequenced the samples and analyzed the data together with the previous dataset using the same service provider. The results showed that the relative abundance of Streptococcus and Veillonella increased in the final samples from both allergy subjects. The ratios of change varied between subjects and genera (Figure 1D). Therefore, the results support my theory that a low abundance of oral probiotics is the cause of allergies. The differences between the NSA23 and NSA25 samples from the non-allergic family members were likely due to an unrelated cause (Figure 1 D).
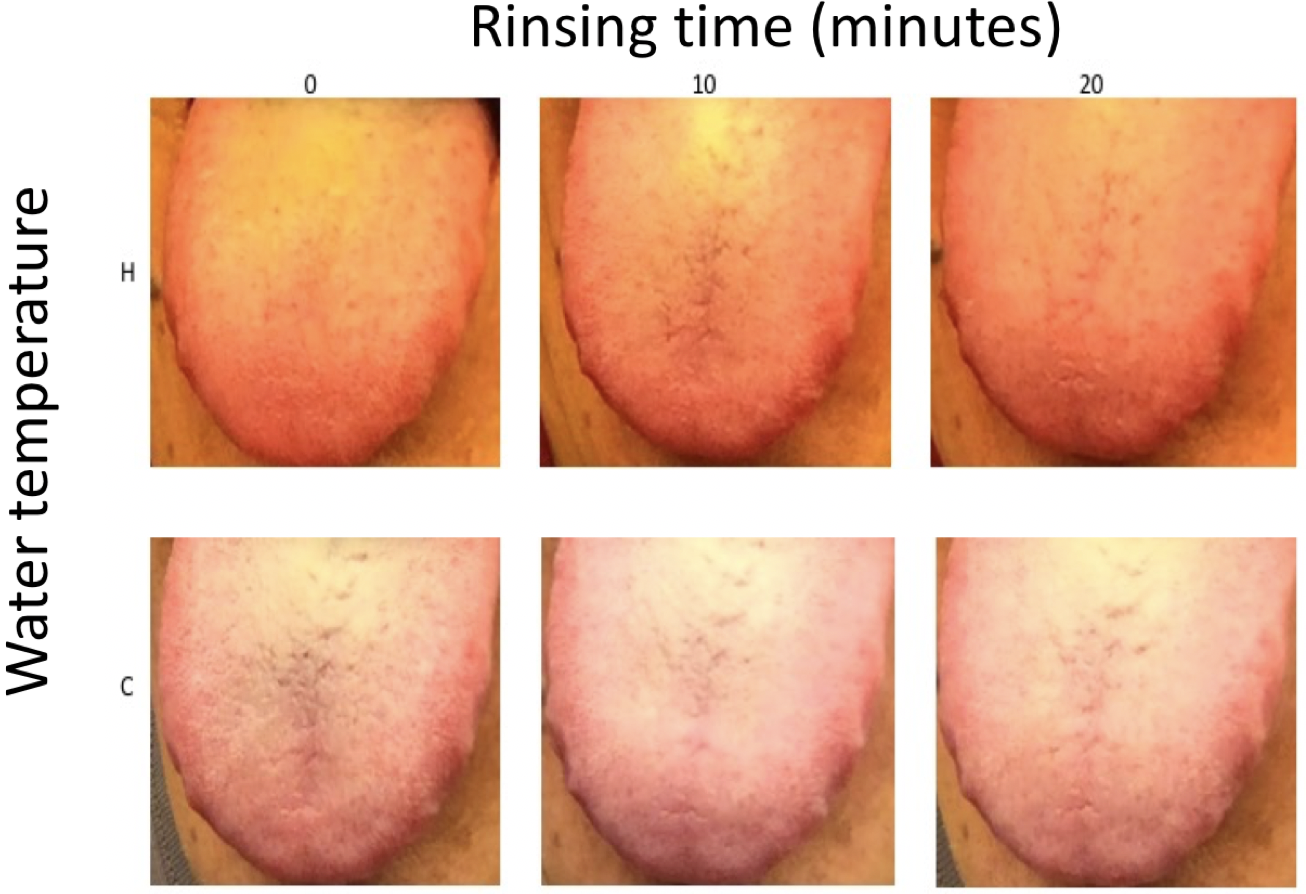
Cross-sample diversity analysis confirms that prebiotic treatment likely caused structural changes in the oral microbial community. Sample CSA25 after the prebiotic treatment significantly differs from the samples collected before the prebiotic treatment (Figure 2). The samples after the treatment were closer to those from the non-allergic family members (Supplementary Figure 2). These changes in oral microbiota are consistent with our theory on the cause of allergies.
The induced microbiota changes and immediate and lasting clinical responses to the intervention, though from a limited sample size, suggest a new theory for the etiology of allergy rhinitis — the Theory of Negative Trigger (TNT) — in which probiotics are the negative triggers that unleash the power of the immune system.
TNT recognizes that the local immune system next to the respiratory tract is at the root of common allergies. This local influence is greater than that from the immune system in the gut. Metabolites, including SCFA(Short Chain Fatty Acids), produced by local beneficial bacteria, likely act as a continuous messenger/pacifier to the nearby immune system. Modern life events, such as extreme oral hygiene or antibiotic usage, local pyrotherapy, or a fever, can remove/diminish trigger bacteria (Figure 3).
Under healthy conditions, responsible probiotics produce enough pacifying metabolites so that the immune system does not react to commensal bacteria and environmental allergens that do no harm (Figure 3A).

Figure 3B demonstrates how common allergies develop. Intensive oral hygiene causes the biofilm to become vulnerable and collapse under the assault of antibiotics (Abeles et al., 2016). This process suppresses oral probiotics. Species of bacteria that are less sensitive to antibiotics take over the ecospace previously occupied by beneficial probiotics. Without enough pacifying messengers from probiotics, the immune system becomes hostile to allergens. I propose the new name, Oral Probiotic Deficiency, for not having enough good bacteria in the mouth. In this condition, reduced oral probiotics cannot produce enough messages to calm the immune system. The consequences of Oral Probiotic Deficiency may include allergy and autoimmune conditions. In other words, the insufficiency of oral probiotics is the real cause of allergies.
In addition to allergies, a hypersensitive immune system may cause other inflammations in the airway by attacking commensal bacteria (Figure 3B). These can trigger, for example, periodontitis, rhinosinusitis, and tonsillitis. Therefore, this theory can explain previously observed associations between oral “infectious” diseases and common allergies (Arbes and Matsui, 2011). Taking the prebiotic composition relieved the allergic family member’s tonsillitis.
Figure 3C illustrates how the immune system regains its strength under pathogenic infection. First, the invading pathogen directly out-competes probiotics that produce pacifying metabolites. Second, severe infection induces pyrogens that raise body temperature to inhibit or kill probiotics. Some probiotics, including Veillonella, are temperature-sensitive (Carlier, 2015). Consequently, suppressing probiotics will cease or significantly reduce the production of relevant metabolites. That means restraints on, or brake of, the immune system disappear or lose. Third, the pathogen causes damage to normal tissue and releases toxins that stimulate the immune system directly. This stimulation acts like an accelerator to the immune system. The immune system, without the brake and with the accelerator, will then regain its strength to fight pathogens. This mechanism could explain how fevers increase immunity and help the body fight diseases, even cancers (Atkinson, 1979).
Theoretically, too many immune pacifying metabolites from overgrowing probiotics may dampen the local immune system too much. The immune system cannot fend off even the normal commensal bacteria when it happens. As a result, it leads to chronic or acute infection, which would reshape the whole community structure (Figure 3D). Studies on people under everyday life conditions may help verify that prediction. The TNT hypothesis would benefit from further testing from many different perspectives: infection, allergy, and autoimmunity.
First, removing and reapplying the triggers can be relatively easy and quick. Theoretically, the effects of the negative trigger should not last long after removing it. Many different cells in the human body can absorb and metabolize SCFAs. Otherwise, the immune system’s power cannot be released in time to protect the host. Several observations support this proposition. The epidemic of common allergies progresses pretty quickly in a short time at a population level. Oral biofilm disappears in less than a day under moderate fever. The prebiotic compound promotes oral probiotics and produces fast and lasting allergy relief.
Second, the gut has a less significant role in common allergies. This study indicates the interactions between probiotics and the immune system come primarily at the local level. Long-distance interaction or circulation of immune cells likely plays a secondary role.
Third, immune system programming is less critical in this situation. This study showed that allergies starting before two years of age can be reversed with the prebiotic mix. This result indicates that immune system programming may be less important in allergy development. The concept of a critical time for immune system development should be reevaluated to determine what is programmed and what is impacted if critical time is missed.
This is all good news for many who have allergies, autoimmune diseases, inflammations, and other conditions, including cancer. Many people would benefit from the power of the immune system being suppressed. Developing a method to fine-tune the immune system’s strength temporally and spatially under these conditions would improve people’s health. For example, one may activate the immune system by suppressing probiotics in the airway and gut with physical and chemical (including antibiotic) means and by adding immune stimulants (such as a vaccine). This may help to control chronic infections and cancers.
The TNT theory also suggests the first critical component of a microbiota associated with a host should be communication with the host. Bacteria in this role should first not cause immediate harm to the host and second should be able to send pacifying signals to the host immune system. This communication is likely achieved in humans through bacteria producing short-chain fatty acids. Streptococcus and Veillonella are in the mouth and airway, fiber-digesting bacteria in the gut, and C. Acnes on the skin. It is not known how widely this mechanism exists across animals/plants. The second component of a healthy microbiota should be sufficient diversity to occupy all nutrient space with members not harmful to the host. The last component is guest members dropped in accidentally.
In short, we have discovered that the cause of many allergies is an oral probiotic deficiency, which I have verified with longitudinal, cross-sectional, and translational studies. Our new TNT suggests possible solutions to many practical issues. It explains why it is beneficial for parents to transfer their microbiota to their children by confinement of mother and newborn at the beginning of life, and fever should be kept if it is not too high. This theory also suggests what comprises healthy microbiota and whether it is possible to reshape microbiota for optimal immune status.
Ethics approval and consent to participate: all participants consented to participate.
Consent for publication: N/A
Availability of data and material: The sequence data is available upon request.
Competing interests: CSH owns Knoze Jr Corp., which holds granted/pending patents covering the oral microbiota-inducing method described.
Funding: N/A
Author’s contributions: CSH is the sole author and is responsible for the content of this manuscript.
Acknowledgments: I thank my family for giving their saliva samples. This study would have been impossible without their contribution. I am grateful for language editing by Shena Han and James Kennedy III. I thank Armand Dichosa and Joe Alcock for reading and commenting on the manuscript.
